Article Text

Abstract
The recent discussions about alternative sources of human embryonic stem cells (White Paper of the US President’s Council on Bioethics, 2005), while stirring new interest in the developmental potential of the various abnormal embryos or constructs proposed as such sources, also raise questions about the potential of the derived embryonic stem cells. The data on the developmental potential of embryonic stem cells that seem relevant for ethical considerations and aspects of patentability are discussed. Particular attention is paid to the meaning of “totipotency, omnipotency and pluripotency” as illustrated by a comparison of the developmental potential of three-dimensional clusters of blastomeres (morula), embryonic stem cells, somatic or (adult) stem cells or other somatic (non-stem) cells. This paper focuses on embryoid bodies and on direct cloning by tetraploid complementation. Usage and patenting of these cells cannot be considered to be ethically sound as long as totipotency and tetraploid complementability of embryonic stem cells are not excluded for the specific cell line in question. Testing this poses an ethical problem in itself and needs to be discussed in the future.
- EMT, epithelial–mesenchymal transition
- SSC, somatic stem cell
Statistics from Altmetric.com
The continuing debates about the ethical aspects of research on human embryonic stem cells are primarily nourished by the unsolved problem that early embryos need to be sacrificed to produce such cell lines, and that, across all traditions and beliefs, people are divided on the question of whether the promise of considerable therapeutic breakthroughs can be weighed against the ethical problem connected with the destruction of early human life.1,2 Embryo destruction is, however, not the only ethical problem that must be seen here. As pointed out earlier, although mostly not discussed in the broader public, the developmental potential that is a basic characteristic of embryonic stem cells must be seen as an additional problem of its own and should even be considered the more challenging problem.3–5 It becomes clear when reading carefully through the recently published White Paper of The President’s Council on Bioethics,6 that this is probably even more complex and more difficult to deal with than the problem of embryo destruction.
The aim of the White Paper is to discuss publicly several alternative procedures that may be chosen to produce human embryonic stem cells, thus circumventing and avoiding the use of normal human embryos (potential human beings who could otherwise have been allowed to develop after embryo transfer to a uterus). The various new sources for embryonic stem cell production that are discussed in the White Paper are not the main topic of this article.i
The point that I would like to discuss here is that this White Paper, in contemplating the ethical aspects of these various procedures, clearly asks for a clarification and evaluation of the developmental potential of the constructs in every case (artefacts, atypical or pathological embryos or embryo surrogates). Should there be reason to believe, after such evaluation, that these alternative sources of cells or their descendants have to be considered to be developmentally equivalent to a normal human embryo, the authors of the White Paper would see an ethical problem (citation: “… ethically unproblematic and acceptable for use in humans … provided the line between pluripotency and totipotency can be maintained” 6, p 59). Interestingly, it seems that this view is in agreement with the arguments mentioned above3–5 and calls for a careful and detailed discussion of the aspect of developmental potentiality of the cells we are dealing with. As we will see, this will lead to conclusions on ethical acceptability and patentability that are somewhat different from what the White Paper proposes.
POTENTIALITY AS APPLIED TO MORULAE AND VARIOUS OTHER TYPES OF CELL CLUSTERS
In the public ethics debate on the use of early human embryos, the inner cell mass (embryoblast) of a blastocyst (the usual starting point for the production of embryonic stem cells) is very often termed “just a cluster of cells”, as is a morula. Embryologists, however, are still divided on the question of whether a morula or the inner cell mass consists of a homogeneous population of cells with equal developmental information or whether there is any cryptic spatial information embedded in this cluster. Specifically, the question is whether cell differentiation and embryonic pattern formation (embryoblast v trophoblast, primary endoderm and axes, such as the embryonic–abembryonic axis and later the main body axes) depend simply on the position of cells in the cluster (inside–outside hypothesis), on interaction with the environment (eg, the uterus) or on pre-patterns derived from the zygote (reviewed in Denker7). Recent evidence suggests that a pre-pattern in the sense of developmentally marked asymmetries does exist and that axial information derived from the zygote and maintained through cleavage does have an important role here. Hence there is a need to view morulae and blastocysts as having pre-patterns that are relevant for future embryonic axes.8,9 It seems logical to assume that such axis-specifying pre-patterns (asymmetries) are lost when embryos are disaggregated to form embryonic stem cells and when these cells are continuously disaggregated during subculturing. It is reasonable to ascribe organism wholeness to morale and blast cysts and to assume that this organism city is lost in the course of these procedures (“organism death”, in the terminology of the White Paper6 applied, egg, to defective morale with many degenerating blazoners). The question remains as to whether the ensuing embryonic stem cells have properties that force us to regard them as cells that are nevertheless so close to early embryos that they deserve special protection with regard to potentiality and exceptional treatment on ethical questions and patent ability, which I believe is the case and which I will try to explain later.
We can base our discussion of the ethical aspects of potentiality on certain embryological facts already established and generally accepted. For the sake of simplicity I propose to discuss four different “clusters” of cells:
-
a morula
-
a cluster of embryonic stem cells
-
a cluster of somatic (adult) stem cells (SSCs)
-
a cluster of other somatic (non-stem) cells.
Are there reasons to regard, with respect to potentiality and ethical aspects, a morula as being equally distinct from a colony of embryonic stem cells as from any somatic stem cells or any group of cells in the body?
Before we look more deeply into this matter, we should deal briefly with two principal objections against such a discussion, which can be found in the literature, in particular with respect to totipotency. Some would argue that there is no point whatsoever in discussing these potentialities because the criteria of developmental potential are believed to be of no help in the ethics debate, as any cell nucleus of the body can be considered to be totipotent in the sense that it can support embryonic development if transferred to an (enucleated) oocyte or zygote (eg, Dolly, the sheep).10 We can deal with this objection briefly because it is incorrect. The nuclear transfer type of experiment shows reprogrammation of the nucleus by the egg cytoplasm, but not cell potential. Developmental potential can be ascribed only to whole cells and depends on intricate interactions between the nucleus and cytoplasm. An argument sometimes heard in Germany (specifically making reference to the wording of the German Embryo Protection Law, Embryonenschutzgesetz) is that totipotency is of juridical or ethical relevance only if it is a property of a single cell (not a group of cells).11 I propose to disregard this argument in the context of this discussion, as the real players in most developmental processes in higher animals including humans are cell groups, not single cells.
Returning to the four different clusters of cells mentioned earlier (morula, embryonic stem cells, SSC and non-stem cells), we ask, “Are there any differences between these clusters that may be relevant to potentiality and simultaneously to ethical aspects?” In doing so, we will concentrate on three-dimensional rather than two-dimensional test systems because embryos need to develop complex three-dimensionality (to acquire the ability to lead an independent life as an individual). Many of the inductive interactions that direct cell differentiation also need a three-dimensional cell arrangement (while planar, ie, two-dimensional patterning is more rare). This is worthy of mention because two-dimensional differentiation systems are most often used in embryonic stem cell research (eg, in the second phase of the embryonic stem cell test12). Two-dimensional systems may allow us to detect only the potential of cells to differentiate into various cell types and thus provide criteria for a judgement on pluripotency or omnipotency5 (table 1). Of primary ethical interest, however, is totipotency, defined as the ability to give rise to all cell types of the body (omnipotency) and also to initiate formation of a harmonious three-dimensional embryo that may finally acquire the ability to lead an independent life as an individual (given the proper environment for development). This necessarily involves early embryonic patterning processes culminating in the formation of a basic body plan by gastrulation.7
Proposed terminology: pluripotency, omnipotency and totipotency
To focus on this ethically most relevant question about totipotency, I will therefore restrict the discussion to three-dimensional biological systems of testing:
-
Autonomous development: the teratoma model (in vivo); embryoid bodies (in vitro).
-
Non-autonomous development: tetraploid complementation.
TERATOMA FORMATION
When embryonic stem cells are transplanted to extrauterine (ectopic) sites they form teratomas, a specific type of tumour consisting of derivatives of all three germ layers.13–15 This tumour-forming potential can be a problem with respect to any use of embryonic stem cells in transplantation medicine and has thus received attention because so far it does not seem to be completely manageable.16 By contrast, with a focus on totipotency, this teratoma-forming capacity of embryonic stem cells does not prima facie seem to provide any arguments that can be used in ethical considerations. A mature teratoma consists of various types of cells and tissues, but in a chaotic arrangement lacking the high structural order that is necessary for organismic wholeness and for the ability to lead an independent life as an individual. An impressive degree of self-structuring potential of the cells becomes obvious in teratomas, although not at the organismic level (basic body plan) but at the organ level (eg, formation of gut-like structures with fairly normal histological stratification and even a degree of functional autonomy17,18). Teratoma formation thus does not seem to indicate that any processes of individuation occur in this situation. It is sometimes argued, therefore, that teratoma formation after embryonic stem cell transplantation even shows a lack of totipotency (in the sense of formation of a harmonious embryonic anlage) in embryonic stem cells. This reasoning, however, is logically incorrect: A teratoma develops also from normal embryos if these are transplanted to the same ectopic sites.14,15 This clearly shows that normal early embryonic pattern-formation processes are disturbed at these extrauterine sites. The reason seems to be that early embryonic pattern-formation processes in mammals are very sensitive and need to be protected from, for example, non-physiological asymmetry cues from the environment.7 This is suggested by observations on twinning as well as on the disturbance of early embryonic pattern-formation processes observed when mouse embryos are kept in in vitro culture beyond the implantation stage.7
Teratomas (consisting of derivatives of all three germ layers) are thus formed after transplantation to extrauterine sites by embryonic stem cells and by early embryos; they are not reported to be formed by other (non-pluripotent) stem cells, nor by other somatic cells of the body (table 2). Although this is a remarkable similarity between embryonic stem cells and early embryos that sets them apart from other types of cells, teratoma formation by itself does not provide any arguments for or against any totipotency that these cells may possess.
Summary of the main aspects of developmental potential discussed in this paper
EMBRYOID BODY FORMATION
When embryonic stem cells are cultured in vitro as three-dimensional aggregates (either free floating or only loosely attached to a substratum), they can form morphologically complex structures called embryoid bodies. This term was originally coined specifically for the mouse system in which these structures closely resemble the core structure of a postimplantation embryo morphologically at the egg cylinder stage (for a morphological illustration highlighting the similarities see Denker7,19). Typical embryoid bodies in the mouse consist of epiblast (the primary epithelium which gives rise to the entire body of the embryo proper through formation of the three germ layers, ectoderm, definitive endoderm and mesoderm, at gastrulation), a central cavity (equivalent to the proamniotic cavity) and an outer epithelium (hypoblast/primitive endoderm). Embryoid bodies in the mouse lack trophoblast and parietal endoderm (ie, extraembryonic tissues that make up the main part of the placenta and the yolk sac, respectively), in contrast with embryoid bodies in primates (see later). By extrapolating from the mouse to the human, it has been argued repeatedly by some authors (particularly in Germany) that embryoid bodies in humans cannot be considered to be totipotent structures because trophoblast is needed, for example, for implantation and placentation. This argument cannot be sustained, however, owing to the fact that embryonic stem cells in primates (including human) have a pronounced potential to differentiate trophoblast (in contrast with the embryonic stem cells in the mouse).7,20,21
Primate embryos (non-human and human) have a different morphology from the mouse at equivalent stages, with no egg cylinder but a flat embryonic disc. To be really embryo-like, an embryoid body would, therefore, more or less have to resemble this type of structure and, for purely physical reasons, we would expect it to form with higher probability in flat (although somewhat three-dimensional) cultures than in free-floating spheroidal cell aggregates, as in the mouse. For this reason the fact that formation of embryonic anlagen has not been observed when human embryonic stem cells were cultured as spheroids22 does not provide a fair arguement in the sense that it would demonstrate lack of embryo-forming potential (totipotency). In fact, structures closely resembling embryonic anlagen of early postimplantation stages have been described as forming in flat dense cultures of embryonic stem cells in the marmoset.23 These structures were astonishingly similar to early primate (eg, human) postimplantation stage embryos (for an illustration, see fig 2 in Denker7). Even an amnion and a yolk sac equivalent were found and, most remarkably, in the middle of the epiblast equivalent, a structure with a primitive streak-like morphology was seen and was interpreted as such by those authors. This meant that processes of definitive germ layer formation and individuation seemed to be occurring. Unfortunately, marmoset embryonic stem cells were not available internationally during the subsequent years, and these interesting observations could not be repeated until now; it was only very recently that new embryonic stem cell lines in the marmoset were developed by another group.24
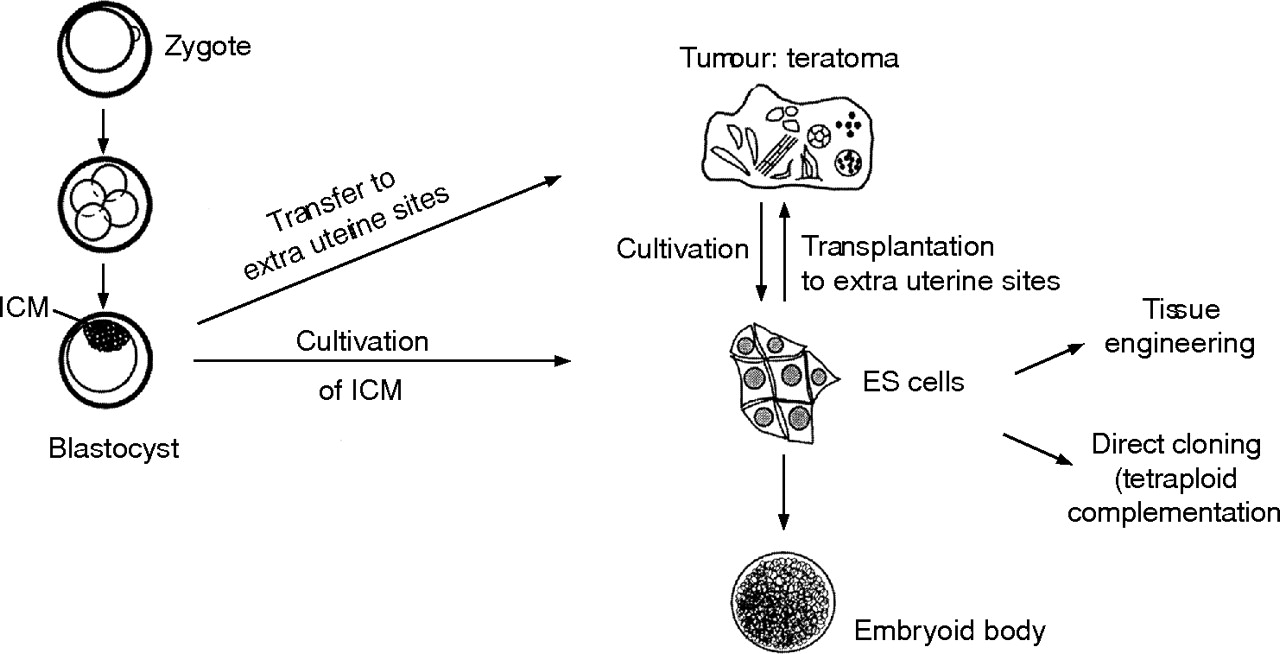
Differentiation potential of embryonic stem (ES) cells. ICM, inner cell mass (modified after Rohwedel et al25).

{kind=link}
{kind=link}
Tetrapoid complementation allows highly efficient direct cloning of normal mice (including a normal germ line, symbolised by sperm) from embryonic stem cell lines in one step. The embryonic stem cells are combined with manipulated (tetrpoid) host cells (blastomeres or, as shown here, a blastocyst) with restricted developmental potential (forming only extraembryonic cell types). Embryonic stem cells show in this case a remarkable developmental potential and form, after transfer to a uterus, the complete body of a mouse without direct participation of the host cells. (Modified from Denker19.)
Literature on embryonic stem cells from other non-human primate species or humans does not report on the formation of differentiated stem cell colonies (or embryoid body equivalents) that would morphologically resemble early postimplantation stage embryos closely. As mentioned earlier, this could be because the three-dimensional culture systems used are mostly not appropriate for formation of a flat embryonic disc as typical for human or primate embryos. Apart from this, it seems possible that differences exist between cell lines with respect to the early embryonic pattern-formation potential.26 This may be because epigenetic peculiarities arise in an unpredictable way and to an uncertain extent during in vitro culture.27,28 In my opinion, it would not be defendable to carry out studies with embryonic stem cell lines in humans with the aim of choosing in vitro culture conditions that may allow these cells to develop early embryonic anlagen comparable to those of the marmoset. I agree with Pera29 that “if such structures were observed in cultures of human ES cells, there would be justified reason for concern, since such an entity might bear a very close resemblance to the embryo near the 14-day limit for observation in vitro”.3,4
My group has recently carried out investigations on differentiating colonies of embryonic stem cell lines in the rhesus monkey (line R366.4). That particular cell line had originally not been reported to form in vitro any embryonic structures resembling postimplantation stages such as embryonic stem cells in the marmoset.30 We basically confirmed this with respect to lack of formation of an amnion or a yolk sac equivalent.31 What we saw, however, was the transient formation of an epithelial disc that often developed a central depression in these colonies, and mostly only one such structure. Here, the epiblast-like embryonic stem cell layer seemed to undergo an epithelial–mesenchymal transition (EMT) as in a primitive streak, accompanied by the typical down regulation of macromolecules and junctions of epithelial cells (E-cadherin, 20-1, tight junctions and the epiblast marker Cx43). In parallel, the colonies markedly increased transcription of Snail2, a central regulator of EMT processes. All of this suggests that what we observed in these colonies is the spontaneous initiation of processes that occur during gastrulation in mammalian embryos. The central depression in these colonies was wide; it was not narrow like a real primitive streak, nor elongated or excentrically located. Hence, these colonies obviously had no positional cues like those that define the anterior–posterior (cranio–caudal) axis of a primate embryonic disc. Also of interest in these observations was that all cells of these colonies (not only those in the central depression) expressed high levels of the genes brachyury and goosecoid. As these two genes are markers for those epiblast cells of an embryo that are in the process of primitive streak and mesoderm formation, we may conclude these particular embryonic stem cells are already triggered from the start of colony formation on to initiate EMT and a gastrulation-like process. However, in the culture system used (which included mouse feeder cells), however, the overall morphology did not come very close to that of a primitive streak—that is, the gastrulation stage of a real embryo.
Gastrulation is of utmost importance not only for formation of the definitive germ layers but also of one of the main body axes, the anterior–posterior axis, and the basic body plan.7 The molecular and cellular biology of gastrulation is the centre of interest of developmental biologists and much progress has been made recently.32 Classical experiments on the germinal disc of the chick and modern molecular approaches in mammals and the zebrafish show that the induction of organiser equivalents and the formation of primitive streaks and the anterior–posterior axis strongly depend on local asymmetries that can be disturbed easily and that respond in a very sensitive way even to non-specific physicochemical signals (reviewed in Denker7). Embryonic stem cells are thought to represent in vitro equivalents of embryonic disc (epiblast) cells and can thus be expected to maintain the ability of sheets of epiblast to respond to external (even stochastically arising) asymmetries by initiating the typical cascade of local gene activation events that lead to organiser and primitive streak formation. From the point of view of developmental biology, we should, therefore, not be too surprised about observations such as those reported by Thomson et al23 on formation of a primitive streak-like structure in marmoset embryonic stem cell cultures, or the EMT processes in those of the rhesus monkey (see earlier). Lewis Wolpert coined the aphorism: “The most important event in your life is not birth, marriage, or death but gastrulation” (Stern32 Preface). The process of twinning (which occurs in humans most frequently at stages after formation of the blastocyst; for literature, see Denker7) may be one example of this. Gastrulation (primitive streak formation) marks (and is instrumental in) individuation. This is the reason for defining the 14th day of development as a limit for embryo experimentation in the UK. As our discussion focuses on ethics, it is relevant that, although embryonic stem cells show a gastrulation capacity, like early embryos, this has never been reported for any clusters of SSC or other somatic cells (table 2).
DIRECT CLONING BY TETRAPLOID COMPLEMENTATION
Embryonic stem cells can form an entire, normal organism when combined with helper cells, either blastomeres from cleavage stages or blastocysts, that have been made tetraploid.33,34 This is a variant of the chimera formation technology, and the use of tetraploid (rather than normal diploid) helper embryos or cells restricts them to forming only extraembryonic cell types (such as placental trophoblast) and not contributing to the formation of the body of the resulting mouse. Tetraploid complementation is a technology that allows for cloning of mice in a one-step procedure and is being used worldwide in many laboratories. We have no reason to doubt that tetraploid complementation would also work with embryonic stem cells in humans if attempted. Although experiments in the mouse show that the efficiency of cloning by tetraploid complementation depends on the particular mouse strain from which the embryonic stem cells were derived, we must, by extrapolation, regard embryonic stem cells in humans basically as potential human beings as long as it has not been shown that the respective cell line cannot form an embryo by tetraploid complementation. The ethical dilemma is evident. Although there seems to be consensus worldwide at this moment that reproductive cloning of humans should be banned, opinions differ considerably on research cloning. Interestingly, Buddhist authorities are expressing a view that is quite opposite to what dominates in the Western world. Depending on the time point of ensoulment that they envisage, they would ban embryo destruction by therapeutic cloning but not reproductive cloning.35 This alone shows that we cannot be sure about any worldwide consensus on the ban of reproductive cloning.
As embryonic stem cells in humans must thus be regarded as potential human beings (even though the efficiency of embryo formation by tetraploid complementation remains unproved for the individual cell line), they must also be considered principally non-patentable.36,37 This may be different for different embryonic stem cell lines, but testing this by performing a tetraploid complementation cannot be an ethically acceptable way. The same considerations must apply for the biological “artifacts” proposed by Hurlbut38(see also The President’s Council on Bioethics6) and any embryonic stem cell lines derived from them.
What embryonic stem cells show when tetraploid complementation is carried out is of course aided (not autonomous) development. The exact role of the helper cells or embryos has not been clarified. They obviously provide for this construct trophoblast cells that are needed for implantation in the uterus, but they may also provide the necessary asymmetry cues for a regular formation of the body axes (gastrulation, primitive streak; for a discussion, see Denker7). We can argue whether tetraploid complementability shows totipotency or not (eg, only omnipotency; table 1). This will depend on whether we feel that it is sufficient for the definition of totipotency that the cells form the whole body of a normal individual including the germ line or whether we want to include the ability to also form the extraembryonic structures autonomously (placenta and yolk sac). (It should be noted in passing that non-human primate and human embryonic stem cells, in contrast with those in the mouse, can differentiate trophoblast by themselves5,7). No matter how we may want to decide about this terminology, it seems to be a peculiarity of embryonic stem cells as compared with other cell types that they show this embryo-forming potential by tetraploid complementation. They seem to share this remarkable potential only with early embryonic cells but not with other cell types, including somatic or adult stem cells. No report exists in the literature about successful tetraploid complementation with somatic stem cells. Somatic stem cells can form chimeras when combined with normal (but not tetraploid) embryos and they then show their plasticity by contributing to the formation of various organs and tissues (the high plasticity reported in Clarke et al39 has been debated by other groups40 and seems to depend considerably on the cell type). The formation of a whole embryo or living animal from somatic stem cells by tetraploid complementation has not been reported in the literature and does not seem to be possible. The developmental potential of embryonic stem cells must thus be seen as categorically different from that of somatic stem cells or other somatic (non-stem) cells (table 2).
CONCLUSION
Embryonic stem cells, being embryoblast-like or epiblast-like cells, show a remarkable developmental potential that exceeds what is usually discussed (ie, the ability to form different cell types: pluripotency or omnipotency). They also have a potential for early embryonic pattern formation—that is, morphogenesis from an amorphous cell cluster by gastrulation to a basic body plan. This potential is shown to only a limited degree in vitro when the culturing conditions restrict orderly pattern-formation processes. (This is seen in the same way with normal embryos of implantation/postimplantation stages.) The fact that embryoid bodies are usually not much more than a caricature of normal embryos does not argue against the totipotency (in the sense of pattern-formation potential) of embryonic stem cells. Nevertheless, remarkable pattern-formation processes by embryonic stem cells in vitro have been shown in the marmoset 23 and, in a more limited way, in a rhesus monkey cell line.31 Far too little is known about embryonic stem cell lines in humans, possibly because these cell lines are usually not allowed to form differentiated flat colonies (as would be physiological for this species). Assisted development (with tetraploid helper cells added) shows, however, that at least mouse embryonic stem cells can form complete normal individuals, consisting exclusively of embryonic stem cell progeny, and this most probably also applies to human embryonic stem cells. This remarkable potential seems to be a peculiarity of embryonic stem cells (table 2), which puts them close to embryonic cells up to the epiblast stage. The (at least theoretical) possibility of cloning human beings from embryonic stem cells in the one-step procedure of tetraploid complementation needs to be put into ethical considerations. It certainly must be seen as an obstacle for patenting human embryonic stem cells as long as the possibility of embryo formation by tetraploid complementation has not been disproved for the human embryonic stem cell line in question.36 As embryonic stem cells can be propagated indefinitely and can thus be distributed worldwide and grow for long periods, the risk that they could theoretically also be used for reproductive cloning of human beings in an unforeseeable future must be included in the information given to patients when informed consent is obtained in the course of embryo donation.37
When embryonic stem cells are compared with early embryos, with other (somatic/adult) stem cells or non-stem cells as regards totipotency (defined as the potential to form a viable individual), a remarkable property that embryonic stem cells are seen to have is their potential to develop a basic body plan and to acquire organismic wholeness if appropriate conditions are provided, and they seem to share this only with early embryos or the embryoblast or epiblast (primitive ectoderm) of those. These remarkable and peculiar properties should force us with respect to ethical considerations to regard embryonic stem cells as categorically, not just gradually, different from somatic stem cells or non-stem cells. Single embryonic stem cell lines may differ with respect to totipotency so defined. Biological tests of totipotency are ethically non-acceptable in humans (this would be reproductive or at least research cloning). With these aspects in mind, the use of any embryonic stem cell line in humans that is suspect of totipotency must be considered to be ethically problematical, and these cells must be regarded as non-patentable. Specifically, the ability to form a normal individual via tetraploid complementation (tetraploid complementability), although not showing autonomous totipotency but a sort of assisted development, must be regarded as problematical under ethical aspects and should indeed preclude patentability. These considerations must apply to any type of stem cells suspect of totipotency, no matter by what procedures they have been derived (including, eg, the “organismically dead embryos” as well as “biological artifacts”, discussed in the White Paper of the US President’s Council on Bioethics, 2005).6
Acknowledgments
This article is based on a lecture given at the workshop Going to the roots of the stem cell controversy, Norwegian Center for Stem Cell Research, Oslo, Norway 13–14 June 2005. I thank the recent and former members of my department who contributed to our own stem cell work, in particular Rüdiger Behr, Bärbel Gobs-Hevelke, Carola Heneweer, Hans-Peter Hohn, Birgit Maranca-Hüwel, Gudrun Mikus, Iris von Recklinghausen, Uwe Scheperjans, Dorothee Schünke, Michael Thie and my wife Ulrike Denker.
REFERENCES
Supplementary materials
This is the corrected PDF.
Files in this Data Supplement:
Footnotes
-
↵i (comprising (i) “organismically dead embryos”, 6–8-celled IVF embryos that ceased dividing; (ii) blastomeres obtained by non-harmful biopsy of living embryos; (iii) “biological artifacts”, genetically modified cells/embryos lacking certain properties needed for early development or implantation (eg, a gene that is necessary for trophoblast differentiation), but which may later be reactivated after embryonic stem cell production (method: eg, “altered nuclear transfer” to an oocyte); (iv) reprogramming human somatic cells, for example with the aid of special cytoplasmic factors obtained from oocytes).
-
This work was supported by Kompetenznetzwerk Stammzellforschung NRW.
-
Competing interests: None.
Linked Articles
- Correction
Other content recommended for you
- Stem cells, embryos, and the environment: a context for both science and ethics
- Regenerative medicine: stem cells and the science of monstrosity
- Current Science of Regenerative Medicine with Stem Cells
- A natural stem cell therapy? How novel findings and biotechnology clarify the ethics of stem cell research
- What’s in a name? Embryos, entities, and ANTities in the stem cell debate
- Stem cell therapy for myocardial repair
- The new stem cell biology: something for everyone
- Islet and stem cell transplantation for treating diabetes
- The “future like ours” argument and human embryonic stem cell research
- The moral value of induced pluripotent stem cells: a Japanese bioethics perspective on human embryo research